MANAGEMENT IMPLICATIONS OF RESEARCH INTO CHIRONOMID ECOLOGY
Glowworms are living organisms and as such one of their requirements for life is food. So what do they eat? One answer to this question is chironomids.
This paper is titled "Management implications of research into chironomid ecology" and it is based on the findings of the research I carried out at Waitomo from 1983-84 on the ecology of the Chironomidae of the Waitomo Stream.
As a background to the paper and so you can gain a better understanding of it, I would like to first outline the reasons for my study and what it entailed.
Firstly, you may wonder exactly what chironomids are. They are small flying insects, commonly referred to as non-biting midges as the adults have no mouthparts so cannot feed, unlike the mosquitos and sandflies we all know so well.
These midges belong to the order Diptera which includes all of the 2 winged flies.
These midges provide the main food source for the glowworms at Waitomo (Edwards, 1924; Gatenby, 1959) along with mosquitos, caddis flies, mayflies, stoneflies, sandflies and other small insects (Richards, 1960) (see Figure 1).
The distribution of glowworms inside the cave is almost certainly controlled by the availability of food insects, and food supply probably also controls the speed of glowworm development (Pugsley, 1980). A plentiful food supply in spring/early summer may initiate early pupation and emergence of the adult glowworms. At Waitomo, most stages of glowworm development are present all year but there is an underlying seasonal cycle with a new generation starting late winter/spring each year (Pugsley, 1980). Chironomid midges are therefore an important consideration in the life cycle of the glowworm and were one of the last areas of glowworm ecology that had not been studied. This was therefore the basis of my study and why I undertook this project.
So you can see exactly where these midges fit into the life cycle of a glowworm, I will tell you a little about the life cycle of a chironomid midge. This will also help explain why careful consideration must be given to the management of the area surrounding the Waitomo Caves and the Waitomo Stream itself.
Much variation exists between the life cycles of different midges, depending on temperature and other factors, so I will use for this example one of the larger midges found in the Waitomo Stream (Chironomus zealandicus) (see Figure 2).
Firstly, the eggs. These are laid by the female adult in one gelatinous mass. The number of eggs laid varies but this species usually produces between 300 and 1600 eggs in one mass. These eggs will hatch into larvae after about 4 days (Forsyth, 1971).
There are 4 "instars" or stages in the larval phase and the greatest percentage of the life cycle is spent as a larvae.
The 1st larval stage is free living until it finds a suitable site to construct a tube on or in the sediment. It will stay in this tube until it develops into a 2nd stage larvae when it makes a new tube in which it will live for its 3rd and 4th stages also (Forsyth, 1971). Some species will use only one tube for all larval stages.
These tubes are usually open ended and made of fine organic and inorganic matter, held together by salivary secretions (Forsyth, 1971).
Tubes may be made on the surface of the sediment or they may be made in the sediment and "U" shaped. Body movements are used to create a current through the tube which brings food with it. This food consists of very fine particles of organic and inorganic matter and it is trapped by the salivary "net" at the front of the tube. This "net" is periodically eaten, along with the food it contains, and a new net made (Forsyth, 1971).
One sub-family (Tanypodinae) of midges do not construct tubes at all and are free-living while others construct tubes on vegetation, undersides of leaves, and on stones (Oliver, 1971).
Once the larval stage is over, the change from larvae to adult occurs, also in a tube. During this stage, gases are formed beneath the skin of the pupae and this helps the pupae rise to the surface after 24-48 hours in this stage (Forsyth, 1971).
The adult then emerges at the water surface in a matter of minutes and are able to fly immediately (Forsyth, 1971) and will often fly downwind (Pugsley, 1980). Mating occurs about 24 hours later (Forsyth, 1971).
Some of you may have observed the characteristic swarms of midges that occur at dusk and in the early morning. These swarms are almost entirely made up of males to which the females are attracted. As a female enters the swarm she is intercepted by a male, mating occurs and they drop to the ground. This lasts several seconds after which time the male returns to the swarm and the female rests before laying the eggs.
Adults will live for 2-4 days only as they have no mouth parts so cannot feed (Forsyth, 1971). The whole life cycle varies with temperature but is between 20 (22°C) to 35 (20°C) days long (Forsyth, 1971).
Chironomids fit into the glowworm life cycle in their short-lived adult stage. Adult midges are strongly attracted to the light of the glowworms where they become entangled in the glowworms "fishing lines" (see Figure 3). The glowworms then kill their prey with their jaws and suck out the blood or they will chop up and eat the whole insect when food is scarce (Stringer, 1967).
As you will realise, there are a number of stages in the midge life cycle, so to study these midges and their role as a food source for the glowworms, I carried out 3 types of sampling of the midge population of the Waitomo Stream.
- Sampling of the sediment of the Waitomo Stream at 5 sites, starting at a site near the top caves of Ruakuri and Aranui and finishing at a site within the grotto itself (see Fig. 4). I sampled these sites monthly for 13 months using an Ekman grab. These samples were then analysed in the laboratory to see what species were present and at what times of the year.
- Secondly, I collected samples of the water and the associated small insect larvae, including chironomid midge larvae, that drift into the grotto over a 24 hour period. I did this monthly for 12 months, collecting samples every 3-4 hours over each 24 period. This gave me an indication of what species were entering the cave via the drift and also the variation over a day and over a year.
- My final type of sampling was light trapping. This involved using an ultraviolet light to attract adult insects to it where they fell into a funnel and then a container beneath the funnel. This sampling was done for 12 hours overnight at 3 sites, once a month for 12 months (see Figure 5). This gave me an idea of what adult insects were present inside and outside the cave and at what times of the year.
I will not show you a lot of data at this stage but I will briefly summarise my findings very generally, with respect to each of the 3 types of sampling.
- In the samples taken from the bottom of the Waitomo Stream, that is, the larvae samples, each site was shown to have its own specific mixture of species and population dynamics over the year - there was very little similarity between sites. This can be related to some extent to the conditions at each site, which I will discuss later. It was clear however, that some of the smaller midge larvae species were living associated with the vegetation within the stream and at sites with slightly larger sediment. The largest larvae were dominant in the very fine, silty sediment such as in the grotto.
- My drift sampling showed that only 2 groups of midges dominated the drift. One of these was a small type of larvae, often found within areas of vegetation in the stream, while the other type of larvae was a larger species.
- The adult light trapping showed that different species were present inside and outside the cave and also abundant at different times of the year.
That is a very brief summary but highlights some of my most important findings that relate to the management of the stream catchment and the stream itself and I will be discussing these in more detail later.
In my discussion of the stream in relation to the glowworms food supply, I would like to concentrate on the factors that influence the numbers of larvae living in the sediment of the stream. This is the main stage of the life cycle of a chironomid midge that is likely to be affected by any changes in the environment and area around the stream. Any small alteration to the area around the stream has the potential to have serious effects on the animals in the stream. The adult population of midges available to the glowworms as a food source is the end result of the population of larvae in the sediment and the factors affecting them.
The main factors affecting the distribution of organisms within a stream are availability of food, nature of the sediments, current flow and temperature (Hynes, 1970). Three of these factors can be influenced directly by the management of a catchment and the 4th factor can be affected to some extent (Table 1).
Firstly, I will discuss the availability of food. Many studies have emphasised that plant material from outside the stream, such as riverbank tree leaves, is the most important food source for the small animals living in a stream (Hynes, 1970) and it is made available to other animals in the stream through the food chain e.g. insect larvae are eaten by larger insects who are in turn eaten by fish.
Chironomids have a variety of feeding habits but the majority eat small plants e.g. algae, small animals and detritus (Oliver, 1971).
Midge larvae living in tubes eat mainly algae (which are very small plants) by collecting, grazing, or trapping them in their salivary nets (Graham, 1976).
Other species eat moss and small plants growing on stones. Drift matter, associated bacteria, plankton and washed up small organisms are also important as food for insect larvae (Hynes, 1970). Studies have shown that high biomass or organisms and a high food supply in the sediment are related (Graham, 1976) so food clearly is significant in affecting larval numbers.
The productive capacity of a valley is actually more important than the production of plants in the stream itself as this is providing much of the food for the stream inhabitants (Hynes, 1970).
The original vegetation of the Waitomo area was dense evergreen forest, dominated by rimu, tawa and kahikatea in a swampy valley bottom. Today about 75% of the valley is in exotic grassland and much is used for agricultural purposes (Pugsley, 1980).
Therefore over the years there must have been a considerable reduction in the amount of leaf matter entering the stream. This material is providing not only a food source for midge larvae and other stream organisms that ultimately are a food source for the glowworms - it also provides a habitat for some of the midges who live on the undersides of leaves and on plant stems.
This leaf matter, as mentioned earlier, is fed right through the food chain and is an essential part of the functioning of the whole stream community. Therefore, in relation to the management of the Waitomo area, any suggestion of removing stream bank vegetation from the Waitomo valley should not be considered unless absolutely necessary. This may well apply to your own areas, and is a very important factor.
The amount of streambank vegetation also influences the temperature of the stream, as it affects how much sunlight is able to reach the stream surface and heat the water. This may alter the conditions organisms are adapted to, with adverse effects on the fauna of the stream.
Another advantage of retaining vegetation not only on the streambanks but throughout the catchment is the much more well known attribute of vegetation to stabilise the land and prevent erosion (Table 2). This relates to the 2nd factor that affects the distribution of organisms within the stream - the substrate/sediment or bottom material of the stream.
Much of the catchment at Waitomo is steep to very steep hill country but is stable under its present cover of vegetation. Any removal of this cover could result in serious erosion problems, increasing the loading of sediment into the Waitomo Stream further.
However, the Waikato Valley Authority are well aware of this problem and are actively involved in an extensive revegetation and land protection programme in the Waitomo catchment, which they will be discussing later. Their programme will reduce erosion and sedimentation in the valley, the latter of which is estimated to have increased 5-fold since European settlement (Waikato Valley Authority, 1979), a very significant increase.
Major roadworks in the 1970s were also responsible for a major peak in deposition of sediment in the Waitomo Stream.
Streambank erosion is probably minimal except during high floods, but may also make some contribution to the sediment load of the stream. Planting of trees on streambanks would not only stabilise them but act as a food source for the stream inhabitants, as mentioned earlier.
The primary concern of the siltation occurring in the Waitomo catchment has been the siltation of the Glowworm Cave and the effect the silt has on causing boat access problems. The effects on the stream fauna are usually of secondary concern.
There are a number of side effects that can result from the silting process including the blockage of passages and subsequent flooding, leading to changes in the physical and chemical processes occurring in the cave. This in turn can lead to changes in the climatic conditions in the cave affecting the biology of the cave. Periodic desilting of the Waitomo Cave is carried out to maintain boat access and prevent any such blockages in the cave. Although it has been suggested that removal of silt may remove potential food for the glowworms thus reducing their numbers (Pugsley, 1980), I believe the effects of this desilting can be minimised if it is timed to correspond with periods of low numbers of food insect larvae in the sediment.
Removal of the large exposed banks of silt in the glowworm grotto could in fact enlarge the present area available for insect larvae to colonise and thereby increase the glowworm food supply further.
This is an example of how an essential management activity of a cave, that is usually carried out with little thought given to biological factors, if given some careful consideration, can be carried out with minimal effects on the fauna of the stream and may even be beneficial. As to other biological effects of silting, it can lead to the smothering of bottom areas of the cave, generally smothering the aquatic organisms and/or reducing the oxygen available to them. Conversely, during periods of flooding, scouring of the bottom by the stream sediment load can result. Flooding is beneficial however in that it brings more insect larvae into the cave as well as bringing more food e.g. leaf matter for these larvae.
The mixture of silt and other particles that enter the stream, and also the processes that occur in the stream, ultimately determines the type of sediment present for the insect larvae to live in. This is particularly important in influencing what species are able to live where in the stream. The type of sediment inhabited by chironomids varies considerably between different species. In the grotto and just outside, 2 groups of midge larvae are dominant (i.e. sites 4 and 5). One of these is a large larvae that is typically found in the top 3-5cm of soft, fine bottom sediments that are rich in organic matter (Robb, 1966) so it is in ideal conditions at these sites. However, it is also large as an adult so is only able to provide a food source for the larger glowworms.
As you move upstream however, there are changes in the sediment and the nature of the waterway.
At the swamp, site 2, several types of larvae are again dominant. Although the sampling at this site was done non-quantitatively with a hand net, it was shown to contain a number of different species of chironomid larvae as well as many other aquatic insects. It is probably an important source of insects for the Waitomo Stream which is adjacent to it.
At the remaining 2 sites in the stream, the upstream sites 1 and 3, there was a mixture of species at both these sites - much more diverse than the other sites. These two sites also had the highest recorded numbers of larvae throughout the year, with peaks occurring in January-February.
Both these sites had beds of an oxygen weed (Elodea canadensis) in the stream and this vegetation was sampled separately at site 1. It was found to be a habitat for two species of small midge larvae.
This instream vegetation is clearly important as a habitat for many smaller chironomid larvae who as adults are especially important to the smaller glowworms as a food source.
The vegetation in the stream also provides oxygen to the stream, stabilises the sediment and traps silt.
The sediment sampling brought out a number of points which can lead to management recommendations being made:
It was apparent that the sites furtherest upstream had the greatest diversity and numbers of midge larvae, especially smaller larvae which were associated in particular with debris and instream vegetation. These sites also had a larger sediment particle size than the grotto site. Studies overseas have come up with similar results showing that smaller larvae tend to dominate within areas of vegetation (Sandberg, 1969) and larger larvae tend to dominate outside such areas.
These findings suggest instream vegetation should be strongly encouraged
Clearly, the conditions upstream are particularly important and should be maintained as they must act as a source of chironomid larvae for areas further downstream. Encouraging these conditions downstream, if possible, would also be of benefit to the glowworm food supply.
The development of a swampy area upstream of the cave has been suggested. I believe that such an area would create conditions particularly suitable for the smaller species of chironomid midge, as well as a wide variety of other freshwater insects.
The creation of such an area upstream does rely on one phenomenon to enable it to be of value to the glowworms as a source of food insects and that is drift.
The sampling of the drift that I carried out in this study showed that many of the insect larvae found in the drift were probably coming from outside the cave, drifting in, settling in the sediment in the grotto and developing, over time, into adults.
As there is no evidence that chironomids mate or lay eggs in the grotto itself, it is clear that the drift is extremely important in bringing larvae into the cave where they can complete their development. The nature of the cave, i.e. its shape, changes from a narrow channel to a large, open grotto, must aid the settling of larvae in the cave itself, as well as larval food such as leaf matter that enters in the drift also and is similarly very important.
Drift is a very important part of the stream system and should be maintained and nothing allowed to impede it in any way. Although I did not measure the distance larvae drifted, overseas work has shown that some aquatic larvae are able to drift in the vicinity of 50-60m (Waters, 1972) and probably more, so areas upstream of the cave are important and should be maintained as near as possible to ideal conditions. Insects further than 60m upstream of the cave are also important as a food source for the glowworms as there is often a general tendency for downstream movement of such organisms and they may provide the basis of a new generation downstream that in turn may drift into the cave.
Flooding often increases drift although this was not always apparent in my study but this can be a beneficial effect of flooding. It did however alter the composition of species drifting to include more smaller larvae, again beneficial for the glowworms ultimately.
The final area I would like to discuss is the importance of preventing major inputs of chemicals, oil or other harmful substances into a stream. I am sure you are all well aware of this but you may not be aware of the reason for its effect on the organisms in a stream.
Although as a group, chironomid larvae are very tolerant to organic and industrial pollution, it is mainly the larger species that have this ability and the smaller species, most important to the glowworms, are likely to be most affected.
Pollution itself generally results in a reduction in the amount of oxygen in a stream which has a number of effects, but pH changes, increased fine sediment load, heavy metal ions, pesticides or other noxious organic compounds can kill most species directly or create conditions outside their tolerance range.
A direct effect is often evident near a site of a pollutant but more subtle differences further away from the site are often harder to detect. Very polluted conditions usually result in 1-2 tolerant species that are superabundant.
The presence of oxygen in a stream is essential and is usually provided by algae and aquatic plants, but pollutants reduce the oxygen available to the stream system with wide ranging detrimental effects.
I am sure you are well aware of the problems of pollution and know the dangers associated with them. However, it is an important aspect and one worth reminding you of.
In conclusion, I would like to show you a list of management aims (Table 3) that I would consider appropriate for the Waitomo Stream and the surrounding area. These relate to maintaining the glowworm food supply source.
I hope that by using the Waitomo glowworm cave system as an example you will be able to relate this to areas that you should possibly be giving greater consideration to in your own caves.
Any changes in the environment, no matter how small, can have devastating effects on small insects and animals. Whenever a system involves biological organisms, great care must be taken with them.
It is very encouraging to see the degree of interest and thought being given to biological organisms in the Waitomo Cave system, and credit must be given to the Waitomo Caves management.
Such foresight means the future for Waitomo is very promising and all cave systems should be given similar thought, consideration and care.
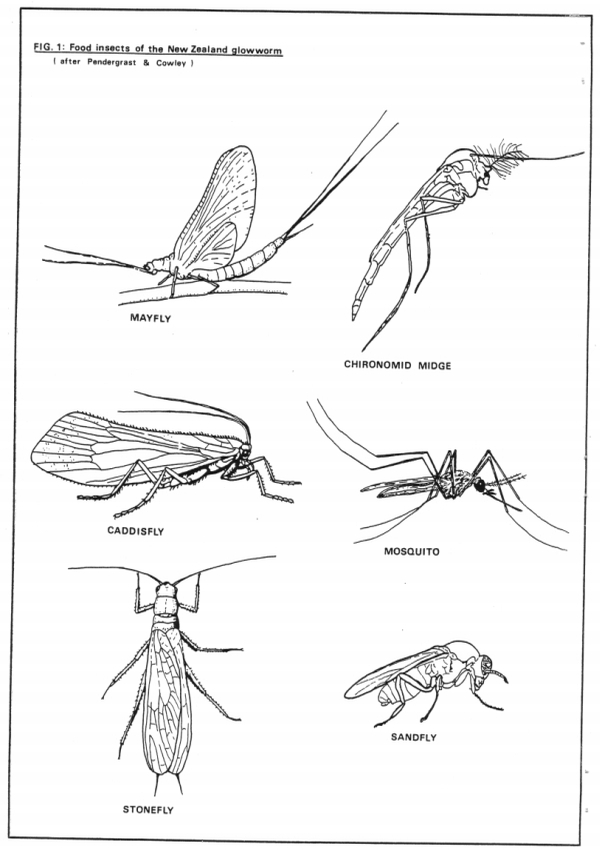
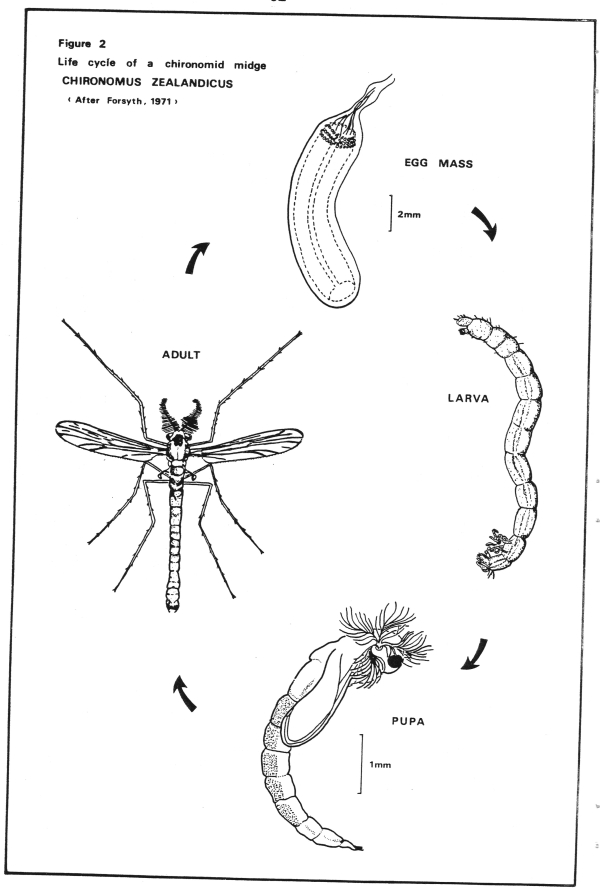
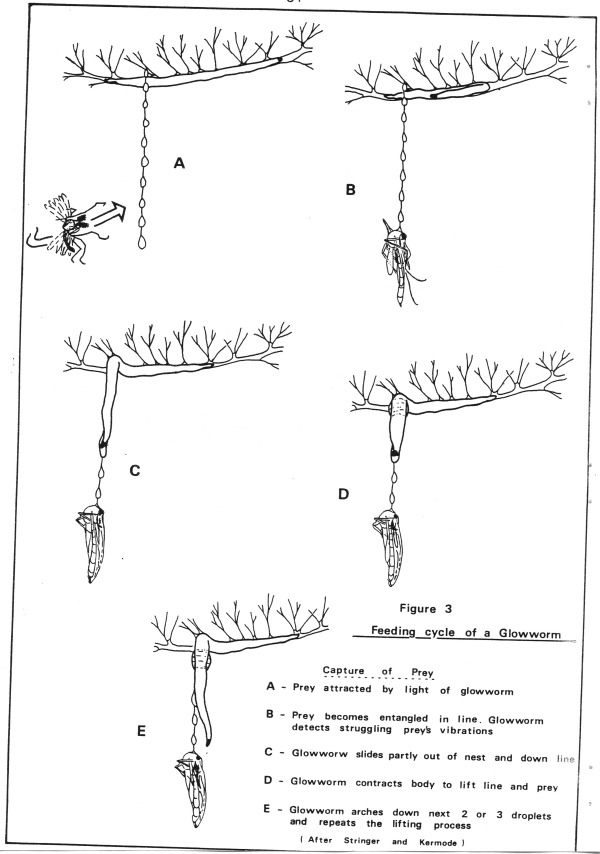
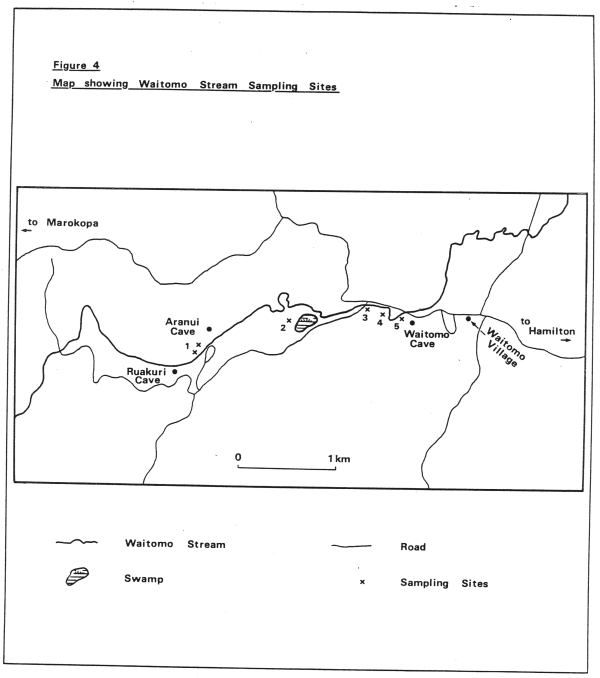
TABLE 1: Factors affecting the distribution of stream inhabitants.
| Sediment | Stream processes |
| Sedimentation into stream e.g. roadworks, erosion | |
| Vegetation in catchment | |
| Food supply | Stream vegetation - source of leaves, detritus |
| Algae | |
| Bacteria | |
| Stream insects | |
| Current | Nature of stream bend and banks |
| Steepness of catchment | |
| Temperature | Degree of shading of the stream by overhanging vegetation affects the water temperature of the stream |
| Thermal pollutants |
TABLE 2: The importance of vegetation in the stream system.
| Streambank vegetation | Reduces bank erosion |
| Provides a food source for stream inhabitants i.e. leaves, detritus. Also part of the drift into the cave | |
| Instream vegetation | Provides a habitat for midge larvae |
| Produces O2 (oxygen) | |
| Stabilises the sediment | |
| Traps silt | |
| Catchment vegetation | Reduces erosion |
| Reduces sedimentation into the stream, cave | |
| Stabilises land |
TABLE 3: Management aims
- MAINTENANCE OF PRESENT FOOD SUPPLY
- Maintaining vegetation within stream and on streambanks.
- Timing management practices to have minimal effect on insect larvae e.g. desilting.
- Retaining swampy areas in catchment.
- Retaining any other conditions favourable to stream life e.g. areas of debris.
- INCREASING PRESENT FOOD SUPPLY
- Development of suitable habitats for food supply insects e.g. swamp, wetlands area
- Removal of exposed silt banks in grotto.
- Encouraging vegetation within and on the banks of the stream e.g. protecting from stock, planting.
- Reducing silt input into stream by reduction of erosion ie stabilising land in the catchment.
REFERENCES
Edwards, F W 1929: British non-biting midges (Diptera: Chironomidae). Transactions of the Royal Entomological Society of London 77:279-430.
Forsyth, D J 1971: Some New Zealand Chironomidae (Diptera). Journal of the Royal Society of New Zealand 1:113-144.
Gatenby, J B 1960: The New Zealand glowworm. Tuatara 8:86-92.
Graham, A A 1976: Ecology and production of Chironomus zealandicus (Diptera: Chirionominae). Unpublished MSc thesis, University of Otago, Dunedin, New Zealand.
Hynes, H B N 1970: Ecology of stream insects. Annual Review of Entomology 15:25-42.
Kermode, L O 1974: the New Zealand glowworm Arachnocampa luminosa. New Zealand Speleological Bulletin vol. 5(91). pp313-328.
Oliver, D R 1971: Life history of the Chironomidae. Annual Review of Entomology 16:211-230.
Pendergrast, J G; Cowley, D R 1966: An introduction to the freshwater insects of New Zealand. Collins Bros and Co Ltd, Auckland. 100 p.
Pugsley, C W 1980: Ecology of the New Zealand glowworm (Arachnocampa luminosa) in caves at Waitomo. Unpublished PhD thesis, University of Auckland, New Zealand.
Richards, A M 1960: Observations on the New Zealand glowworm Arachnocampa luminosa (Skuse) 1980. Transactions of the Royal Society of New Zealand 88:559-574.
Robb, J A 1966: A study of the influence of selected environmental factors on the egg and larval instars of the midge Chironomus zealandicus Hudson. Unpublished MSc thesis, University of Canterbury, Christchurch, New Zealand.
Sandberg, G 1969: A quantitative study of chironomid distribution and emergence in Lake Erken. Archiv fuer Hydrobiologie Supplement 35(2):119-201.
Stringer, I A N 1967: The larval behaviour of the New Zealand glowworm arachnocampa luminosa. Tane 13:107-117.
Waikato Valley Authority 1979: Waitomo Caves Catchment - Water and Soil Management Scheme. Technical Publication 9. 17 p.
Waters, T F 1972: The drift of stream insects. Annual Review of Entomology 17:253-272.