Glowworms are more diverse than we thought: cave and forest-adapted species in Australia
School of Biological Sciences, The University of Queensland, Qld 4072, Australia.
Email: d.merritt@uq.edu.au
Abstract
Glowworms emit light to attract prey into their webs. They are found in suitable wet caves as well as in forests. In wild caves of Tasmania and New Zealand, glowworm populations (Arachnocampa tasmaniensis and Arachnocampa luminosa, respectively) maintain synchronised rhythmic light output, waxing and waning together in a 24 hour cycle. Here I show how the Tasmanian species (and also probably the New Zealand species) is capable of synchronizing the bioluminescence cycle. In laboratory experiments we exposed a single larva to three others that were on a different cycle. The single larva shifted its time of glowing to match the others over about eight days. This synchronisation capability probably allows the glowworm colonies in caves to glow most brightly all at the same time as a way of attracting more flying insects into their webs. In comparison, the south-eastern Queensland glowworm species can’t synchronise. It seems that the synchronisation ability is present only in the species that have large cave populations.
Introduction
In this paper, I point out the substantial behavioural differences among species of Arachnocampa, the glowworms of Australia and New Zealand. The main finding is that cave-adapted species such as Arachnocampa tasmaniensis synchronise their bioluminescence in caves, a form of cooperation that appears to give the participants an advantage in being able to attract prey more efficiently. In essence, it is an approach known as group foraging.
First, I’ll present some background on glowworms. They are members of the genus Arachnocampa. They produce light to attract prey into their sticky snares. Light (bioluminescence) is produced in cells located at the tips of internal tubular structures branching from the gut, known as Malpighian tubules. In most insects they function solely as excretory structures, but in glowworms they have taken on a dual function; excretion and light production. Arachnocampa glowworms are the only insects that produce light in this way. The light-producing cells are located internally at the posterior end of the larva. The cuticle is transparent to allow the transmission of light. In addition, an internal reflector composed of a mass of air-filled respiration tubes is present around the light-producing cells (Figure 1). In this case, they function both as a respiratory system and a light reflector.

Figure 1: The posterior end of a glowworm larva showing the silvery-white reflector surrounding the light organ.
The distribution and relationships of glowworms are keys to understanding the behavioural and evolutionary differences between species. Claire Baker’s research compared the traits of the New Zealand and one Australian glowworm (Baker and Merritt, 2003) and the morphology and DNA sequence of all known glowworm groups in Australia and New Zealand (Baker et al., 2008). She named a number of new species and – most importantly for this paper – provided a phylogenetic tree: a reconstruction of the most likely relationships among the present-day species, with estimated times (millions of years ago) when the major divergences took place. A few key points are that (1) as expected, the most ancient divergence is between the New Zealand species and all of the Australian species. (2) The Australian species can be divided into two main groups or subgenera: subgenus Campara and subgenus Lucifera (Figure 2). The subgenus Lucifera has only two member species: Arachnocampa tasmaniensis and Arachnocampa buffaloensis. They share the traits of inhabiting cooler regions and having significant cave populations. The subgenus Campara includes the remaining six Australian species, distributed from the wet tropics region of northern Queensland to the cooler temperate forests of the Otway Ranges in Victoria. They are predominately found in forest settings where the habitat is cool, shaded and damp, but some populations of these species are found in caves.
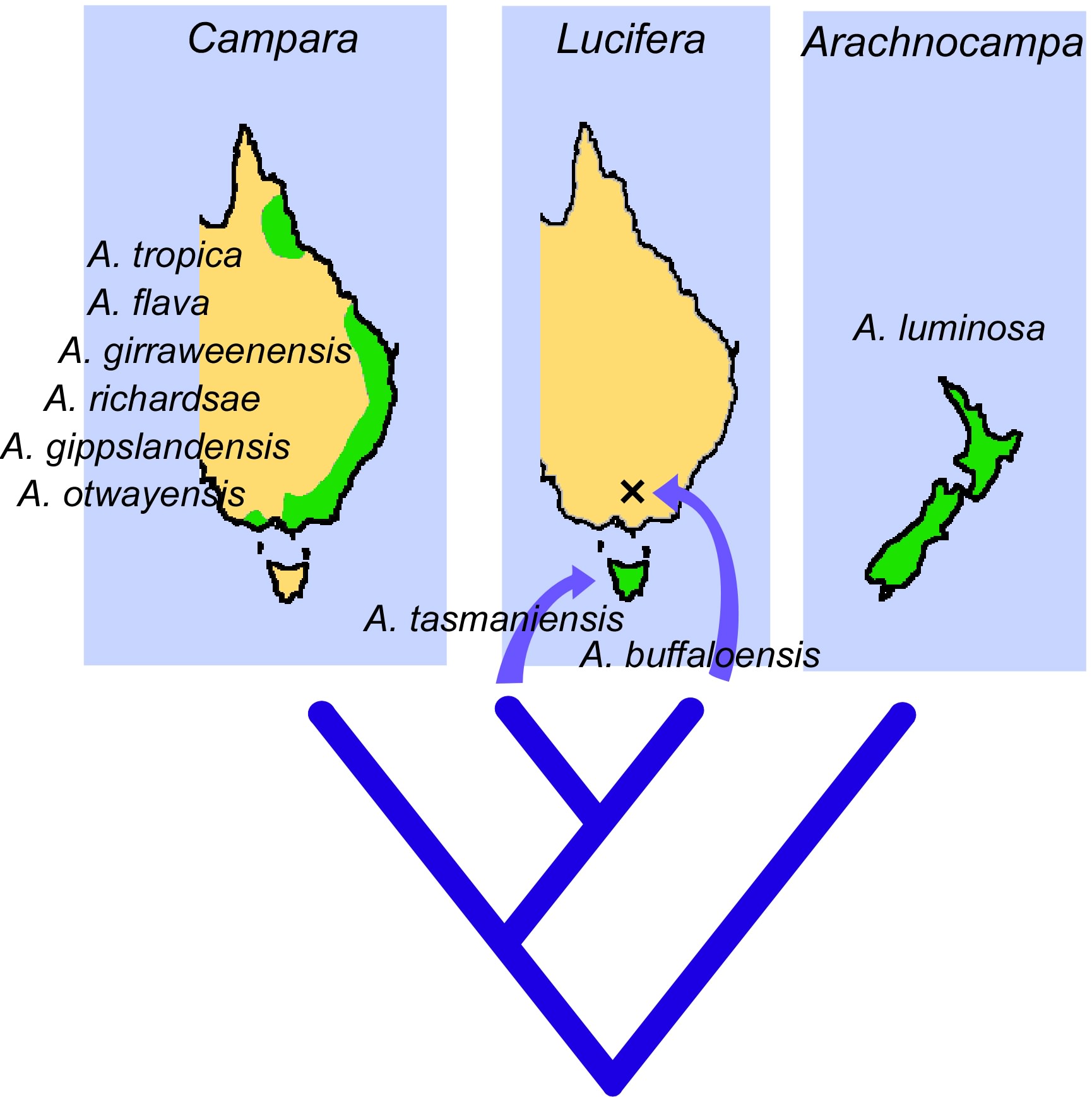
Figure 2: A phylogenetic tree showing the relationships and distributions of the Australian and New Zealand species of glowworms.
Students at the University of Queensland, School of Biological Sciences have been researching bioluminescence regulatory mechanisms. It has been known for some time that extraneous light causes larvae to cease glowing, as noted by some of the pioneering researchers on glowworms (Richards, 1960). This reflex is understandable; there is no advantage in expending energy producing biological light when it is barely visible and cannot compete with non-biological sources. The larvae have eyes so they can see light, although we don’t know how sensitive they are. Larvae in forest habitats will glow throughout the night, modulating the intensity so that it is brightest after dusk and dims toward dawn. A number of factors other than light will cause an increase or decrease in bioluminescence intensity, including vibration and loud sounds. Rainfall causes a large increase in light output.
Because glowworms react to natural light, they have no obvious need for a circadian clock that controls their bioluminescence, but because so many animals show biological rhythms that control behaviours such as sleep-wake cycles, we tested for clock control of bioluminescence in the local forest species, Arachnocampa flava. We found that bioluminescence does come under the control of a biological clock (Merritt and Aotani, 2008). When larvae were placed in constant darkness in the laboratory they continue to cycle their light intensity. After a week or so, the amplitude of the daily cycles had damped, but a persistent rhythm was apparent. The rhythm is known as a circadian rhythm because it doesn’t keep an exact 24 hour periodicity, rather individuals showed different periodicities, usually greater than 24h hours, just as humans do.
In a forest-adapted species such as Arachnocampa flava, the regular waxing and waning of solar light through each 24 h period entrains the clock every night and the presence of the clock control is only obvious under controlled experimental conditions. You may ask why have an endogenous control of an activity such as light production when a reflex-like response to the daily day-night cycles is sufficient to regulate the behaviour? This question is difficult to answer; however, many circadian biologists propose that the endogenous rhythm allows animals’ metabolism to prepare for lights-on or lights-off and buffers them from initiating immediate but inappropriate responses to darker than normal or brighter than normal conditions.
The demonstration of a biological clock that controls the bioluminescence made us curious about what happens in cave populations. Glowworms in the deeper zones of caves never see daylight so it was possible that they would be glowing at a consistent level all the time because their clock – if they have one – has never been entrained by daylight. Alternatively, individuals might keep their internal rhythm of glowing but individuals in a colony would be out of phase with each other because each has its own personal periodicity.
The way to test this was to apply long-exposure, time-lapse photographic methods that would operate in a cave environment. Affordable digital SLR cameras were combined with time-lapse controllers and battery power allowing us to set up a system that would take a photo every 10 minutes while unattended for several days. Afterwards, the images are converted to grey scale and the image analysis software, ImageJ, used to count the number of larvae visibly glowing in each frame, to measure the intensity of each individual and the total intensity of larvae in the field of view over time.
The initial results from Marakoopa Cave in northern Tasmania – since confirmed in other Tasmanian caves – were unexpected. Individuals show precise 24 h cycles in their intensity of glowing. Within a colony individuals are synchronised, i.e. they showed the same time of peak and trough in their intensity curves. Consequently, the intensity of light produced by any single colony oscillates in the form of a sine wave. Some individuals never turn off completely; others dim to the extent that they are no longer detectable by the camera, but they all maintain the same periodicity. Another surprising outcome was that the time of the peak bioluminescence of colonies located well within caves tended to occur in the late afternoon. Colonies in different caves show different times of peak intensity and there seemed to be some plasticity, i.e. it might be earlier or later from one year to the next. Understandably, it is hard to be definite about this because the recording method gives a snapshot of periodicity over a few days at a particular location.
One explanation for the colony-wide cycling is that larvae synchronise to each other’s bioluminescence. If proved, this would be significant because no such bioluminescence synchronisation had been shown in any organism. One comparison is the synchronously flashing fireflies of south-eastern Asia. The male beetles congregate in large numbers in a few trees and synchronise their flashes at about 1 – 2 Hz to attract females. The substantial difference is the fact that glowworm synchronisation is on the scale of 24 hours and involves the circadian system, whereas firefly synchronisation is a higher frequency. We could not eliminate other possibilities, such as the rhythms being entrained by detection of small daily temperature oscillations within the cave. After light, temperature variations are the next most effective entrainment stimuli for the rhythms of many animals.
An in-cave experiment was carried out that pointed to synchronisation but did not prove it. Mystery Creek Cave in Tasmania was chosen because it has a large colony of glowworms and is relatively easy to access. Colonies are present in the cave mouth and deeper. The idea was to focus a light beam on a subset of the main ceiling population to expose them to light for several hours a day for several days (Maynard and Merritt, 2013). The light should shift the phase of their glowing rhythm and, once the artificial light pulses ceased, they should resynchronise to the surrounding colony. A 12 V 20 white LED lamp was focussed with Fresnel lenses to create a 1.6 m diameter spot encompassing 78 larvae on the main ceiling colony in Mystery Creek Cave. A 12 V timer was used to expose larvae in the spot to 3 hours of light per day for 5 days. Over the five days of daily light pulses, the spot-lit cluster of larvae progressively shifted their peak glowing time to match the time of light exposure. Once the light exposures ceased, the larvae then progressively resynchronised to the surrounding population. After nine days when the experimental observations had to be terminated, the treated group had almost re-synchronised to the untreated, surrounding population.
We needed to carry out experiments in the laboratory where environmental conditions could be tightly regulated to prove that behavioural synchronisation was taking place and confirm that A. tasmaniensis has circadian regulation of bioluminescence. With permission from Tasmania’s DPIPWE and assistance from Mike Driessen who has been monitoring the cave glowworms (Driessen, 2010), we collected larvae from the cave and set them up in the laboratory in Brisbane where we can maintain them in incubators at a cool 8°C; the mean annual temperature of Mystery Creek Cave.
First, we established that Arachnocampa tasmaniensis – like their mainland cousins, Arachnocampa flava – have a circadian control of their bioluminescence. Second, we showed that light entrains the rhythm, just as it does in Arachnocampa flava, but it entrains the rhythm to the opposite periodicity (Merritt and Clarke, 2009, 2011; Merritt et al., 2013). Light exposures of several hours per day, over several days, in otherwise dark conditions, caused the larvae to adjust their bioluminescence rhythm to peak when they anticipate the lights to come on. This seems counter-intuitive; however, our explanation is that the species is primarily cave-adapted and the ability to synchronise is an important adaptation. The ability of periodic light cycles to entrain them would cause a dark-zone colony to synchronise; an example of mutual social entrainment. But what happens among the many forest populations of this species? They switch on only at night. We took larvae from the mouth of a nearby cave and showed in the laboratory that indeed their endogenous rhythm tells them to glow most brightly during daylight hours, but the daylight itself over-rides the rhythm and causes them to switch off. At dark, the inhibition is lifted and they switch on their bioluminescence. It is as though the biology of Arachnocampa tasmaniensis is centred on cave habitats where synchronisation is a priority and forest habitats are an afterthought. On the other hand, the rainforest-adapted Arachnocampa flava that doesn’t have cave populations (as far as we know) acts like a typical nocturnal animal; its endogenous rhythms tell it to glow most brightly at night.
These experiments told us that light could entrain the rhythms but we did not yet have proof that individuals can synchronise to each other’s glows. Honours student Andrew Maynard designed laboratory experiments that proved synchronisation does take place. Larvae from Mystery Creek Cave were pre-set to different periodicities by exposing them to light for several hours per day over a number of days: one group’s bioluminescence peaked at around 01:00 h and the other at 13:00 h. Then three larvae from one group were exposed to one larva from the other, placed 7 to 10 cm apart (Figure 3). The single larva changed its periodicity over several nights until its period and phase both matched the other three (Figure 4). As predicted, Arachnocampa flava under identical experimental conditions did not show any signs of phase matching. So these experiments proved that the simultaneous waxing and waning of glowworm light within caves is due to a visually-mediated synchronization of bioluminescence intensity.
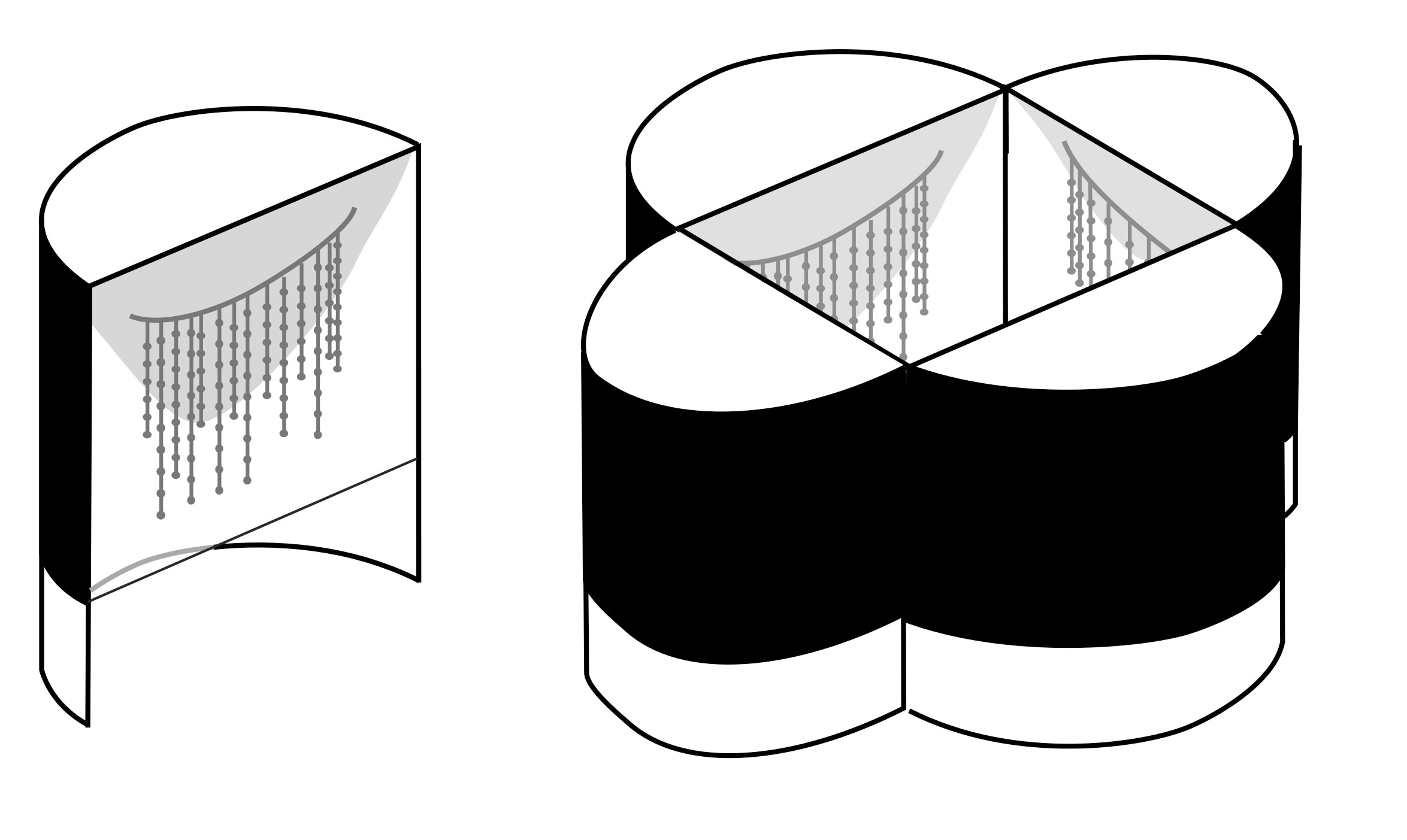
Figure 3: The arrangement of glowworms in individual habitats when they were exposed to each other’s lights.
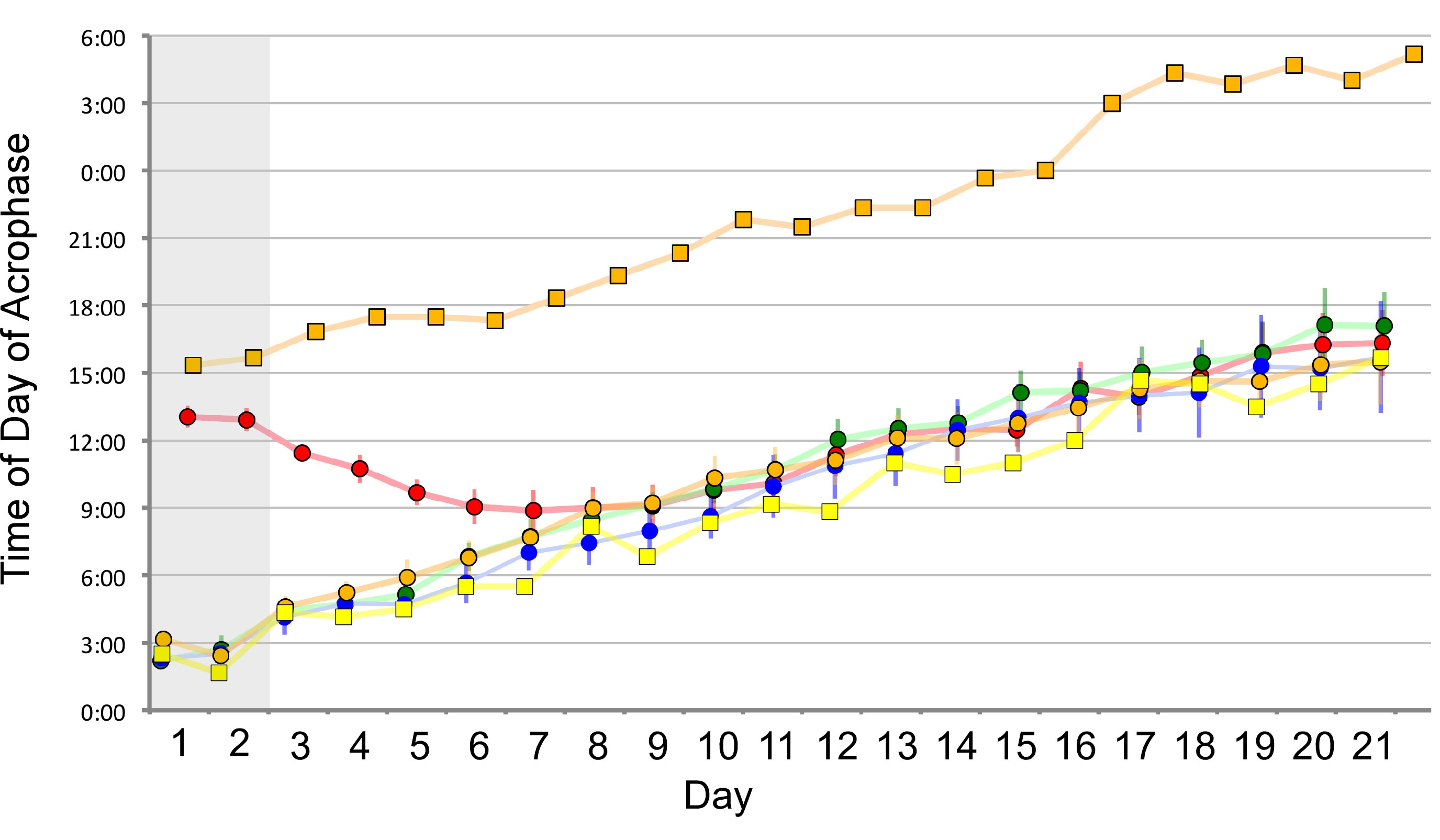
Figure 4: Synchronisation of a single larva (red symbols) to match the glowing cycle of three larvae that were initially synchronised to each other. The orange line is a control larva that was visually isolated from all others.
A second interesting outcome was that under constant darkness the larvae remained synchronised but they did not show the precise 24-hour periodicity that we see in cave populations; rather, they show a period of >24 hours. They entrain to each other but in the absence of any environmental cues the rhythm “free-runs” i.e. the group’s periodicity is the average of each individual’s periodicity and individuals do not have very accurate clocks (remember the definition of circadian means approximately daily). That is why the peak occurred slightly later each day in the laboratory experiments even after all four larvae were synchronised (Figure 4). We suspect that a consistent timing of the availability of prey from day to day could entrain the rhythm in caves, but we haven’t yet done the experiments to test this idea.
Another outcome of the comprehensive photographic sampling was the observation that, while all colonies show the same 24-hour periodicity, they do not all show the same phase. The main ceiling colony in Mystery Creek Cave is composed of many thousands of individuals: it has shown a consistent phase over many years of sampling with the peak intensity occurring at 14:35 h (Merritt and Clarke, 2011). However, another more isolated colony has changed phase between sampling years. Another colony shows even more interesting characteristics. It is located on an outcrop several metres high and wide, encompassing a vertical rock face that transitions to an overhang. The subsection of the colony on the vertical face peaks at 11pm while the adjacent population located on the overhang peaks at 4 am. To explain the different phases we suspect that the subpopulations of the colony can’t see each other due to their location and therefore cannot synchronise. Perhaps the subpopulations attract prey from different locations as well, explaining their different phases. In the future, such explanations can be tested by setting up insect traps inside caves to tell us when the prey insects are most likely to be active. The benefit of our combined approach – controlled laboratory experiments backed up by manipulations and recording in caves – allows us to create a model of how individuals should visually interact and then testing the model inside caves.
Conclusion
It is likely that the New Zealand glowworm, Arachnocampa luminosa, is a synchronising species, based on recording in Hollow Hill Cave and long-term time-lapse imaging at Waitomo Glowworm Cave, but the required laboratory experiments are yet to be carried out. Arachnocampa flava does not synchronise, and preliminary experiments indicate that the other Australian mainland species, Arachnocampa richardsae and Arachnocampa girraweenensis, like Arachnocampa flava cannot synchronise. An aim of future work is to find out which species groups show which behaviour and, if possible, examine the genetic machinery of the biological clock to see if there is any genetic signature associated with the evolution of this substantial change in behaviour among what was thought to be a homogeneous group of related species.
Acknowledgements
I would like to thank Arthur Clarke and Cathie Plowman for their help over the many years we have been working on Tasmanian glowworms. I’d also like to thank Mike Driessen of Tasmania’s DPIPWE for assistance and permissions in working in Tasmania’s world heritage areas. The many students who have worked on the Arachnocampa bioluminescence system at the University of Queensland – Claire Baker, Saki Aotani, Julie-Anne Popple, Rebecca Morley, Robyn Wills, Lisa Rigby, Essie Rodgers, Ami Amir Abdul Nasir, Andrew Maynard – have kept the project running and have all made useful contributions.
References
Baker, C. H., Graham, G. C., Scott, K. D., Cameron, S. L., Yeates, D. K., Merritt, D. J., (2008). Distribution and phylogenetic relationships of Australian glow-worms Arachnocampa (Diptera, Keroplatidae). Molecular Phylogenetics and Evolution 48, 506-514.
Baker, C. H., Merritt, D. J., (2003). Life cycle of an Australian glow-worm Arachnocampa flava Harrison (Diptera: Keroplatidae: Arachnocampinae: Campara). Australian Entomologist 30, 45-55.
Driessen, M. M., (2010). Enhancing conservation of the Tasmanian glow-worm, Arachnocampa tasmaniensis Ferguson (Diptera: Keroplatidae) by monitoring seasonal changes in light displays and life stages. Journal of Insect Conservation 14, 65-75.
Maynard, A. J., Merritt, D. J., (2013). Synchronization of circadian bioluminescence as a group-foraging strategy in cave glowworms. Integrative and Comparative Biology 53, 154-164.
Merritt, D. J., Aotani, S., (2008). Circadian regulation of bioluminescence in the prey-luring glowworm, Arachnocampa flava. Journal of Biological Rhythms 23, 319-329.
Merritt, D. J., Clarke, A. K., (2009). Rhythmic regulation of bioluminescence in glow-worms, Arachnocampa, In: Meyer-Rochow, V.B. (Ed.), Bioluminescence in Focus - A Collection of Illuminating Essays. Research Signpost, Kerala, pp. 291-303.
Merritt, D. J., Clarke, A. K., (2011). Synchronized circadian bioluminescence in cave-dwelling Arachnocampa tasmaniensis (glowworms). Journal of Biological Rhythms 26, 34-43.
Merritt, D .J., Rodgers, E. M., Amir, A. F., Clarke, A. K., (2013). Same temporal niche, opposite rhythmicity: two closely related bioluminescent insects with opposite bioluminesce propensity rhythms. Chronobiology International 29, 1336-1344.
Richards, A. M., (1960). Observations on the New Zealand glow-worm Arachnocampa luminosa (Skuse) 1890. Transactions of the Royal Society of New Zealand 88, 559-574.